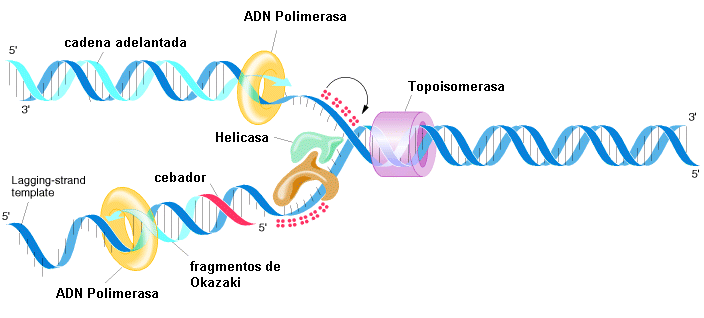
ALTERACIONES DE LA EPIGENÓMICA
Las células eucariotas
requieren mecanismos epigenéticos para controlar la transcripción de los genes
para la especificación y diferenciación celular como:
- · Remodelación de la cromatina
El estado de la estructura
de la cromatina puede ser regulada por complejos de remodelación de la
cromatina dependientes de ATP o modificaciones de las colas de histona. Estos complejos
se pueden clasificar en SWI / SNF, ISWI, remodelación de nucleosoma y complejo
desacetilasa (NuRD), y INO80 sobre la base de sus subunidades ATPasa
catalíticos. La modificación de las colas de la histona es a menudo
enzimáticamente reversible y resulta en una alteración de la interacción entre
la cromatina y el ADN. Estas modificaciones incluyen la acetilación, metilación,
fosforilación, sumoylation y ubiquitinación
- · Metilación del ADN
Se produce típicamente en
sitios CpG que contienen nucleótidos de citosina-guanina en una secuencia
lineal, se encuentran a menudo en los promotores de genes de mamíferos, y el
grado de metilación en estos sitios se correlaciona bien con el estado de la
transcripción de los genes correspondientes. La metilación de ADN posee funciones
para silenciar la transcripción de genes de forma estable

Remodeladores de la
cromatina para el desarrollo cardíaco
Complejo factor asociado Brg1/Brm
Es el complejo SWI / SNF de mamíferos
compuesto de al menos 11 subunidades, y sus variables contribuyen a funciones
distintas durante el desarrollo.
La subunidad ATPasa del
complejo BAF es codifica ya sea por genes homólogos Brg1 (relacionado
Brahma-gen 1) o BRM, Brg1 puede aumentar la accesibilidad del promotor para
factores de transcripción, pero también puede unirse directamente a los
factores de transcripción tales como proteínas Gata para regular la
transcripción de genes. Experimentos en ratones heterocigotos se eliminó Brg1 y
éstos exhibieron defectos morfogenéticos cardiacos.
Polybromo (BAF180) es la
subunidad prominente del complejo PBAF relacionados con BAF, también está
implicado en la cardiogénesis potenciando la activación transcripcional mediada
por receptores nucleares, tales como RXR, VDR, y PPAR. La supresión de Baf180 no
conduce a la letalidad embrionaria temprana pero resulta en una pared cardíaca
muy delgada con una disminución de trabéculas. La ablación de otras subunidades
del complejo BAF (Baf47, Baf155, o Baf250) causan letalidad embrionaria en la
pre-implantación (Baf47, 155) o E6.5 (Baf250) en ratones
WHSC1
Es una histona
metiltransferasa que regula la activación de Nkx2-5, una proteína crítica para
la morfogénesis cardíaca. WHSC1 físicamente se asocia con Nkx2-5 y es
necesario para la regulación negativa de Nkx2-5 y sus genes diana, posiblemente
a través de la trimetilación de la histona H3 en la lisina 36 H3K6me3 causando defectos
del septo ventricular y el tabique en ratones y humanos.
Smyd1 (SET y MYND dominio
1), un miembro de la familia de lisina metiltransferasa, se expresa
específicamente en el tejido muscular y actúa como un represor transcripcional
por catalizar la metilación de histonas a través del dominio SET. En embriones
de ratón con deleción de Smyd1 éstos exhiben defectos cardíacos graves ,
incluyendo la pérdida del ventrículo derecho con la maduración interrumpida de
los cardiomiocitos. En el pez cebra, Smyd1 es esencial para la contracción del
músculo cardíaco y miofibras maduración y sugiere un papel conservado de Smyd1
para el desarrollo cardíaco.
BIBLIOGRAFÍA
http://cardiovascres.oxfordjournals.org/content/91/2/203